Pattern Formation:
Pattern formation is the process of organization of cell in different spatial position according to their identities . Pattern formation play a crucial in forming of tissue and ultimately the organs in thier correct places within the body .
Pattern formation refers to the complex organization of cell with respect to time and space within body . Process of pattern formation controlled by genes . As we learned earlier in previously article about differential gene expression i.e though we having the same content of DNAs in nucleus of cell still it produces different characteristics according to the position and environmental factors which involved in gene expression . A word come in this scenario is ‘ morphogenesis ‘ ( creation of diverse anatomies from similar genes ) .To understand this pattern formation let’s take an example of pattern formation in ‘Drosophila Model‘ (Drosophila melanogaster) .There are three fundamental type of genes which responsible for structure of fruit fly those are maternal effect genes, segmentation genes and homeotic genes . The functions of these genes and thier interaction can well studies by 3 parts i.e
1. Anterior-posterior axis patterning
2. Dorso-ventral axis patterning and
3. Placement of specific tissue in a particular position.
1. Anterior-posterior axis patterning:
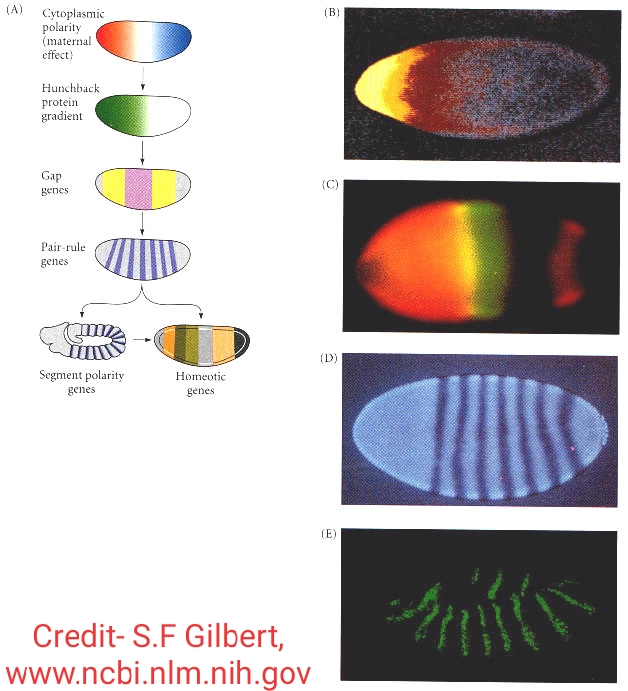
The Anterior-posterior axis development occurs through the polarization which is present in egg . It is generally the cytoplasmic polarity . First of all the maternal effect genes from the ovaries expressed i.e either it repress or activate the expression of certain zygotic genes by diffuse through the blastoderm . Two pairs of proteins produced by maternal effect genes among which ‘Bicoid‘ and ‘Hunchback‘ regulate the production of the anterior structure and another pair of proteins called ‘Nanos‘ and ‘Caudal‘ regulate the production of posterior part of embryo . The zygotic genes in turn by the influence of maternal effect genes the gap genes produces 3 segments whose gene concentration is differ to each part and due to this there is a transcription of paired rule genes occurs and which form 7 segments pattern in anterior-posterior axis which in turn activate the zygotic genes called segments of units . Finally during this transcription gap genes ,pair-rule genes and sement polarity genes combinedly regulate the homeotic genes whose transcription results in the formation of different organs from each segments .
2. Dorso-ventral axis patterning :

Dorso-ventral axis patterning is depends upon gradient of a transcription factor called dorsal .This gradient is a result of cell-to-cell signalling events among such group of cells . The translocation of dorsal proteins from cytoplasm to the nuclei of ventral cell takes place,these translocation process is mainly governed by some sets of maternal effect genes . After the translocation of dorsal proteins done ,the dorsal proteins act upon the cells nuclei and based on different concentrations dorsal protein in the nuclei represents the specific fates of those cells . By the way the mother’s gene is the responsible factor for production of dorsal protein . Dorsal protein is not synthesized untill the message code for translation about 90 minutes after fertilization . After translation the protein is found throughout the embryo still there is a morphogenicity or differentiation in dorsal as well as ventral region found in embryo, it is because of that the dorsal binds to certain genes to activate or supress thier transcription and in turn they get supress because if the dorsal proteins dose not binds with them then they could not be able to specify ventral types and on the other hand by not binding with ventral nuclei the dorsal genes get not repressed and all the cells of the embryo becomes dorsal cells . The mechanism of this translocation followed a cascade process which will be deal in another article .
3. Placement of specific tissue in particular position :
The anterior-posterior and dorso-ventral axes of embryo from a cordinate system which will further define a specific position for placement of specific tissue in particular position . This system follows the ‘Cartesian Model‘ . It is experimentally found that the organ form at the intersection of dorso-ventral and anterior-posterior or region of gene expression .
Cytoplasmic determinant :
During the time development of oocyte in ovary there is a accumulation of certain specialized molecules occurs inside the oocytes which are distributed all over the cytoplasm of oocyte unevenly . These molecules are called as cytoplasmic determinant as during oogenesis in embryo,it plays an important role.
After the multiplication of zygote to became the blastomere ;each and every cell of the blastomere is ascertained by a determinant or groups of determinants . The cytoplasmic determinant distributed among the blastomere at different times depending on the species and type of determinant . Though these determinant has the potential to organ formation therefore the distribution of cytoplasmic determinant in the daughter cells resulting from the first divisions are totipotent in nature and they can independently lead to a complete individual in undifferentiated blastomere .
There are three ways through which a cell become determined for further development .
1. Autonomous cell specification
2. Conditional cell specification
3. Syncytial cell specification
1. Autonomous cell specification :
It is happen during the cleavage of the embryonic development which follows the qualitative separation of zygotic cytoplasm and each cell becomes specified according to the cytoplasmic determinant which they obtained . Each cell is independent from other neighbouring cells interns of cell fate and this type of cell fate is called autonomous cell specification .
Autonomous cell specification give rise to mosaic development of the embryo since the embryo appears to be like a tile mosaic of self differentiating parts . During the mosaic development,the cells of future embryo distinct from each other having the diverse cytoplasmic determinant and which is specific to each cell type .
2. Conditional cell specification :
It is a second way of commiting cell fate where cell undergoes interaction with neighbouring cells . Though these cells have the ability to follow one path of differentiation and the interaction of these cell may affect the fate of one or both the participants . This type of cell fate determination is called conditional specification ,because the fate of the cell depends upon the condition in which they belongs to this type is found in all vertebrates and a few invertebrates .
3. Syncytial cell specification :
It is the third way to determine cell fate ;oftenly observed in many insects and called as syncytial specification . Interaction between the maternal components within the syncytial blastoderm occurs before the cell membrane separating nuclei have formed . Major cell fate decissions are made even before cells have formed . Variable cleavage produces no rigid cell fates for particular nuclei . After completion of cellular process, conditional specification is mostly observed .
Assymetric cell division :
In an assymetric cell division,the daughter cells which are produce having different cellular fates . Remarkable,stem cells divide assymetrically which give rise to two distinct daughter cells i.e one copy of the original stem cells and a second daughter programmed to differentiate into a non-stem cell fate ( en.m.wikipedia.org ) . The process of assymetric cell division was originally described by Ed Conklin in ascidian embryos .
There are two mechanism which cause the distinct properties of daughter cells of dividing mother cells . Suppose daughter cells are initially equivalent but difference is induced among them due to signaling between them from surrounding cells or from precursor cell . This mechanism is known as extrinsic assymetric cell division . For second mechanism i.e the daughter cells are naturally different at the time of division of the mother cell, because this mechanism do not relie on interaction of cell from thier environment or within themselves . Hence it is known as intrinsic assymetric cell division . The term assymetric cell division usually refers to the intrinsic assymetric cell division .
In multicellular organism intrinsic assymetric cell division required two proponents ; 1) the mother cell should be polarized ,2) the mitotic spindle should be aligned with the axis of polarity . Let’s discuss assymetric cell division in three animal models namely – in nematode ( Caenorhabditis elegans),fruit fly (Drosophila melanogaster) and in spiralian .
a) Assymetric cell division in C.elegans:
After the fertilization in the first cleavage of zygote in nematode is a unequal one where one large anterior AB cell and a smaller posterior P1cell produce . This assymetric division shows in the form of anterior-posterior axis . The P1cell acts as a stem cell and on further division gives P-type cells and one daughter cells that follow another path for its development . On the first 3 division of P cells i.e P1 to P2,P2 to P3 and P3 to P4 are responsible as body cells and on fourth cleavage i.e division of P4 cells gives rise to germ cells .
Earlier before fertilization there is no assymetry found in nematode egg . The cleavage pertaining both unequal and assymetry is related to this point of sperm entry, which determined the posterior pole . After first cleavage there is a demarcation of anterior-posterior axis revealed and resulted in large AB cell at anterior pole and small P1 cell at posterior pole occurs the granules called p-granules get located at posterior end and actin microfilament formed a cap at anterior end . On first cleavage the P1 cell retained the p-granules and the presence of microfilament in anterior end dose not control the traditional orientation of cleavage due to some alkaloid (mycotoxin) Cytochalasin D result in the unequal and assymetric cleavage with large AB cell from another end .
A marker called PAR-1 proteins encoded from par-1 gene of maternal genome is the best possible factor for making the anterior -posterior axis,because any disruption in par-1 gene may affects assymetric cleavage though it helps in arranging the microfilament cytoskeleton at posterior end .
b) Assymetric cell division in Drosophila melanogaster :
Maternal gene is the most crucial factor in creating the difference along the axis of egg,even before the fertilization . Generally these maternal which is more expressive in egg do not have any effects on mother’s ovaries .
The expression of these maternal genes can be interprete by observing the effects on larva on mutational basis i.e how the maternal genes cause the mutation in larva . This mutations can be studied into three classes i.e in anterior mutations, posterior region mutation and final one is mutation of both the terminal regions .
Mutation in anterior regions called bicoid mutation leads to reduction or loss of head and thoracic structure , posterior region mutation caused by nanos mutant resulted in loss of abdominal regions and mutation caused by torso mutant at both terminal regions affects specialized structure the acron and telson at head and tail ends of embryo . Each genes acts independently from each other .
c) Assymetric cell division in Spiralian :
The spiralian are morphologically diverse class of protostome animals, including within thier number the molluscs,annelids,platyhelminthes and other taxa . The term spiralia is applied to those phyla that exhibit canonical spiral cleavage (en.m.wipedia.org) .
An example of this asymmetric cell division has been discovered in the mollusc Ilyanasa obsoleta . Polar lobes are formed during the cleavage which contains the material for development of larval shell,foot,operculum,eye etc .
Insitu hybridization and immunofluorescence experiments show that mRNA transcripts co-localize with centrosomes during early cleavage . Consequently these transcripts are inherited in a steriotypical fashion to distinct cells . All the transcripts are followed for body axis patterning and insitu hybridization of transcripts which fails in some other functionality resulted in such localization . However disruption of microtubules polymerization with nocodazole ( tubulin binding agent ) restrict assembly/diassembly of microtubules and of actin polymerization with Cytochalasin B,shows the cytoskeleton is also important in this asymmetry . (en.m.wikipedia.org)
It appears that microtubules are required to recruit the mRNA to the centrosome and that actin is required to attach the centrosome to the cortex . Finally ,introducing multiple centrosome into one cell by inhibiting cytokinesis shows that mRNA dependably localizes on the correct centrosome,suggesting intrinsic difference between each centrosomal composition and this asymmetric divison can be observed after the first two division of the cell